





























Species in the Thripidae exhibit a wide range of biologies. Some, such as those of Dendrothrips and Scirtothrips, breed only on leaves, with Scirtothrips species apparently particularly dependent on actively dividing young tissues for successful breeding. In contrast, many species of Panchaetothripinae breed most readily on old leaves, and often build up large populations on mature leaves of plants that are water stressed. Some Thripidae species breed only in flowers, such as Odontothrips species in Europe and Odontothripiella species in Australia, both of which are associated with Fabaceae, and species in these genera are often host specific. Some flower-living Thripidae have been demonstrated to be effective as pollinators on particular species of plants (Mound, 2004; Zerega et al., 2004).
Many Thripidae feed both in flowers and on leaves, and some of these are the most common pest species and tospovirus vectors on crops. Curiously, a few of these pest species, including Thrips tabaci and Frankliniella occidentalis, may at times act as beneficials, in that they will also feed on spider mites that can be so damaging to plants. A few Thripidae, such as those of the genus Scolothrips, are obligate predators of mites.
The Poaceae, including grasses and bamboos, support a very wide range of Thripidae in a variety of genera. Species of Chirothrips and related taxa breed in the flowers of grasses (and some sedges), and a single individual will pupate within the glumes that would normally surround a mature seed. Other Thripidae, such as Anaphothrips and Stenchaetothrips species, feed on grass leaves, but Limothrips species seem to feed on both leaves and flowers of grasses.
All Thripidae have two larval and two pupal instars before emerging as adults, and pupation usually occurs at ground level (Heming, 1991). Because of their unusual life cycle, Thysanoptera are sometimes referred to as the Remetabola, a sub-group of the Hemimetabola (Heming, 2003).
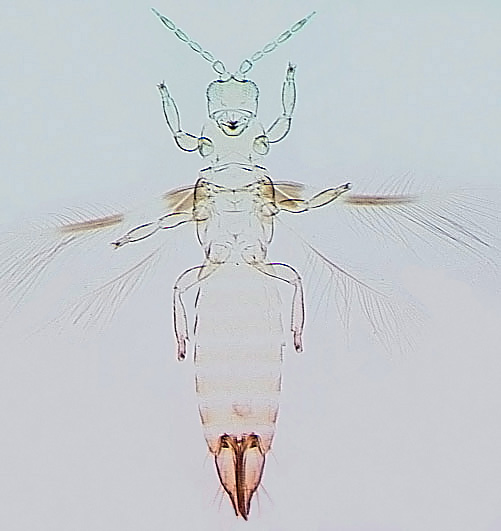
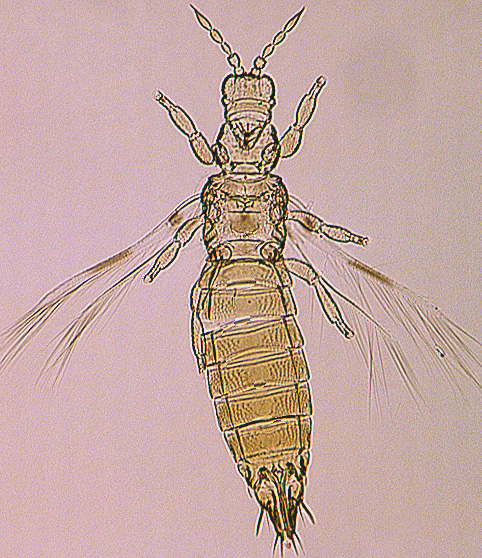
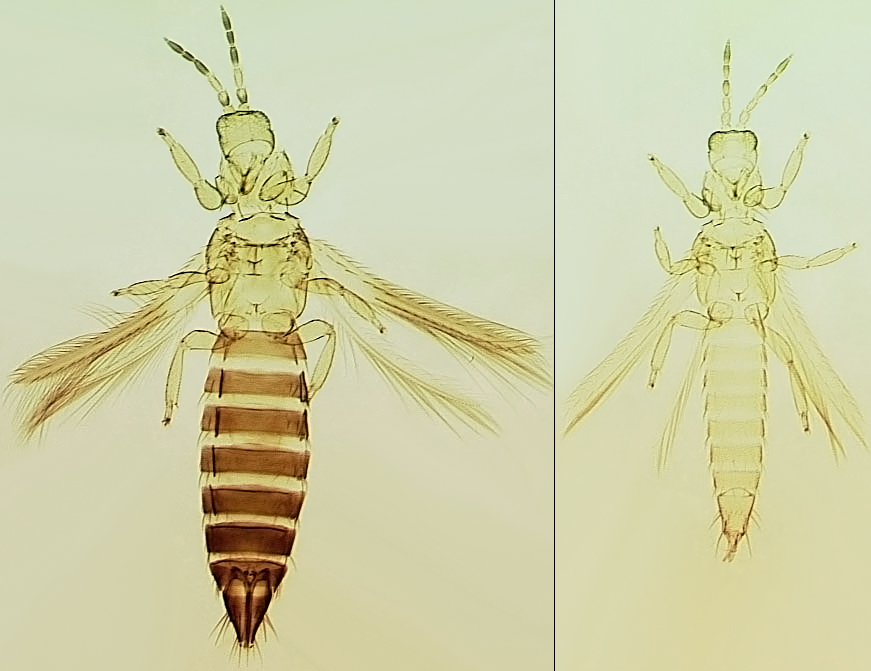
Depending on the species, adults may have fully developed wings, or the wings may be no longer than the thorax width (micropterous), or the wings may not be developed at all (apterous) (see, Tyagi et al., 2008). Males are more commonly micropterous or apterous than females, but species such as Frankliniella fusca regularly produce micropterous females. Winged forms are unknown in many Thripidae, such as the species of Aptinothrips and Apterothrips.
Most species of Thripidae are bisexual, although males usually constitute 30% or less of any population. For a few common species, such as the North American Basswood thrips, Thrips calcaratus, males have never been found, but individual populations of some other thrips, such as Thrips tabaci, commonly lack males.
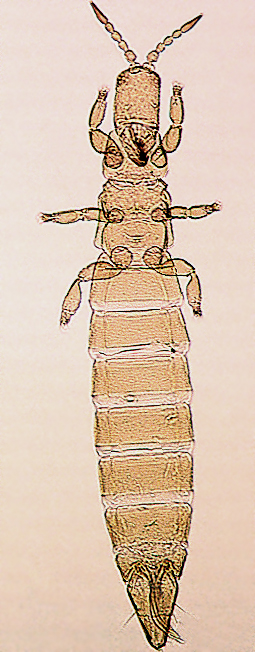
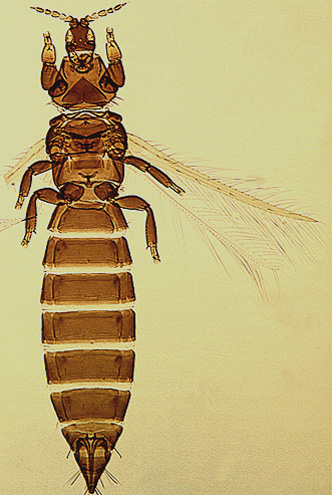
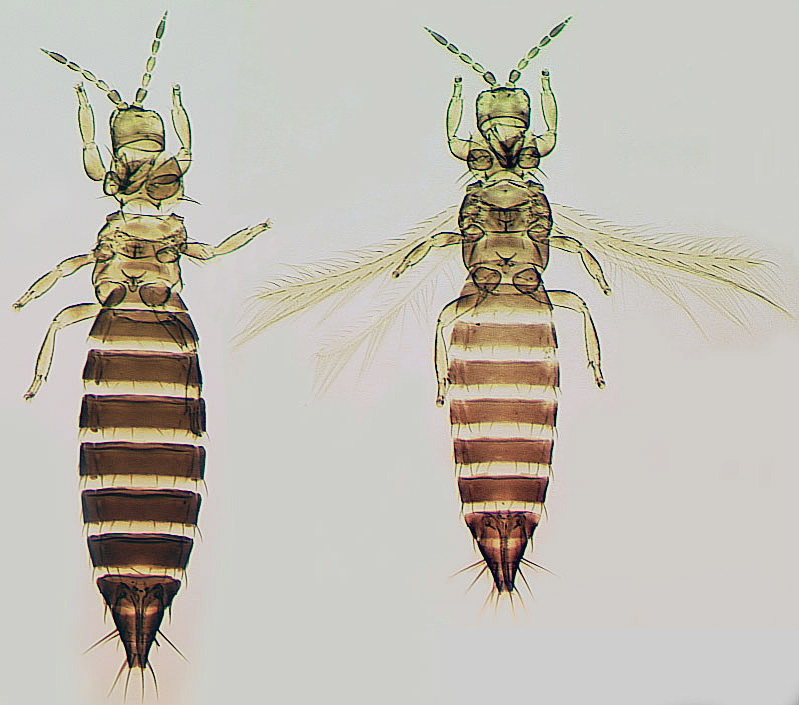
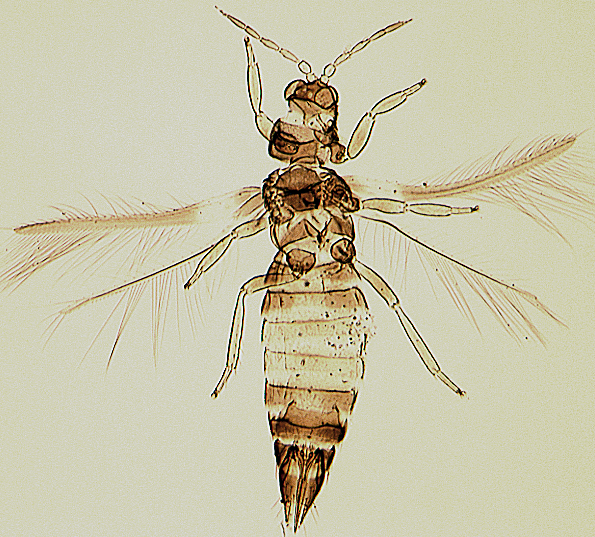
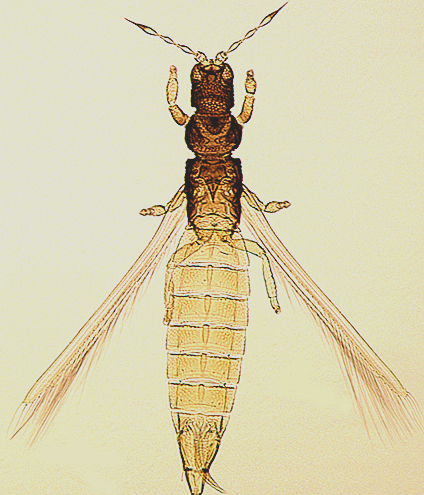
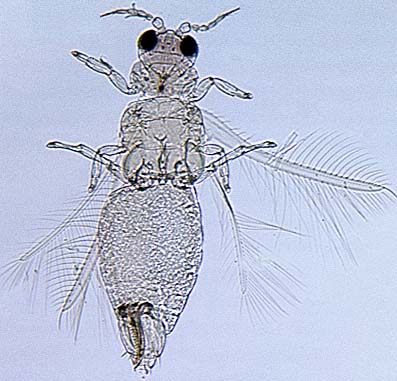
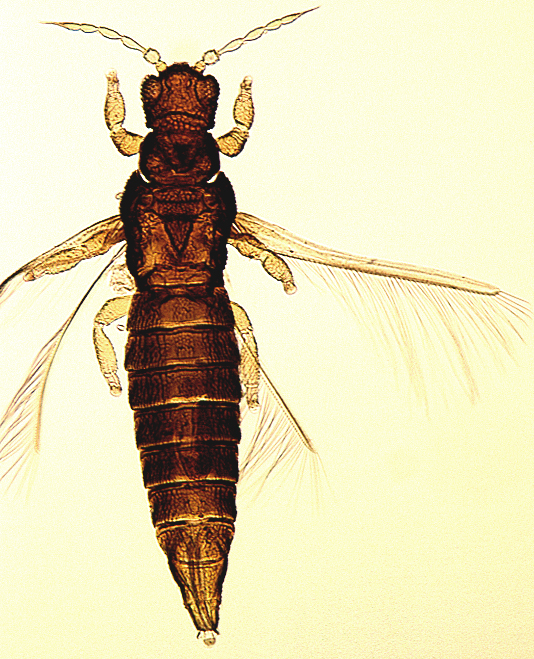
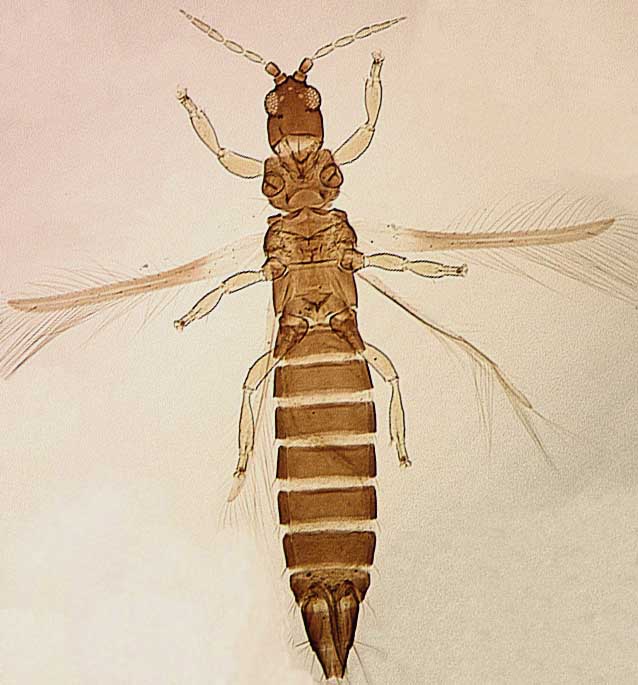
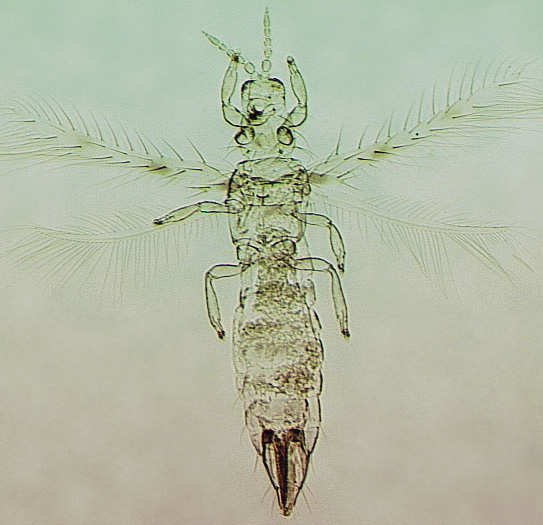
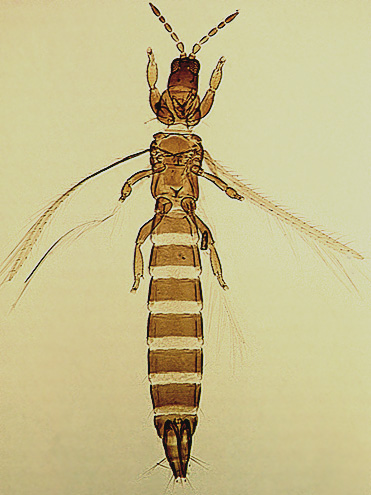
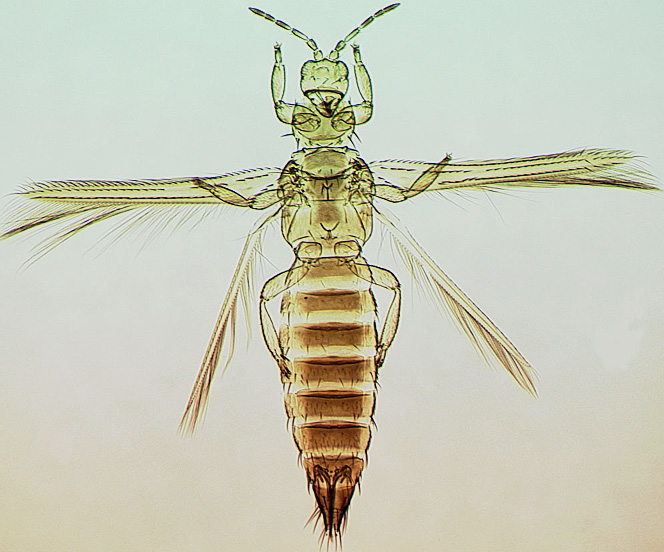
The Thripidae is a diverse family, in colour, body sculpture and thoracic structure, but can usually be recognised from the structure of the antennae and slender forewings.
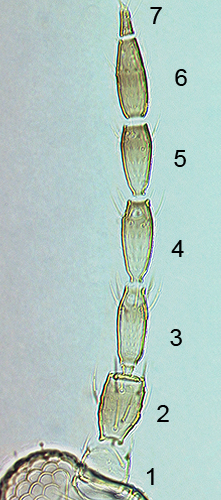
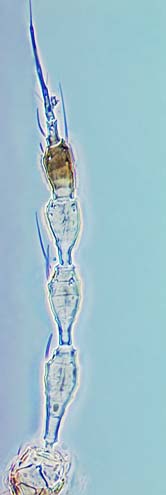
The number of antennal segments varies from six to nine among Thripidae, and the sensoria (trichomes) on segments III and IV are characteristically Y-shaped but sometimes simple. Most Thripidae have the antennae 7-segmented or 8-segmented, and the comparatively rare 9-segmented condition in this family is possibly derived secondarily rather than an inherited ancestral condition.
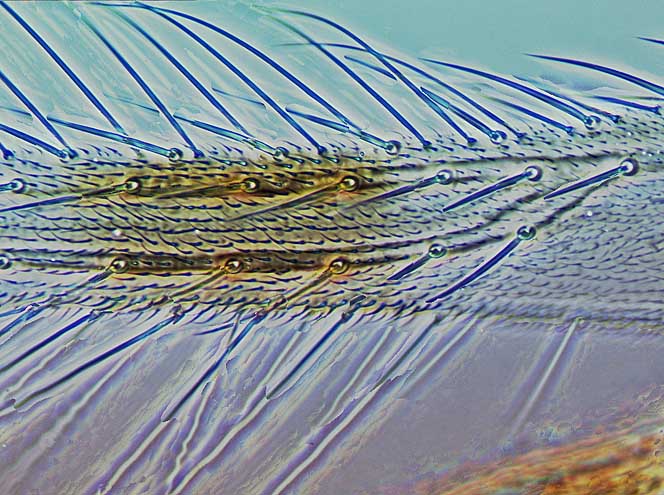
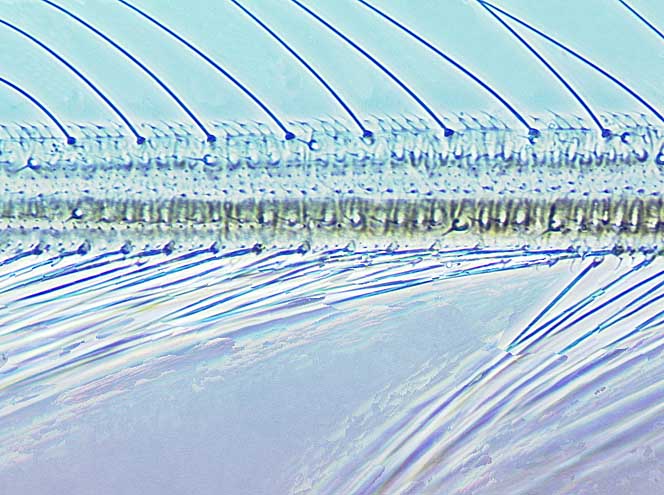
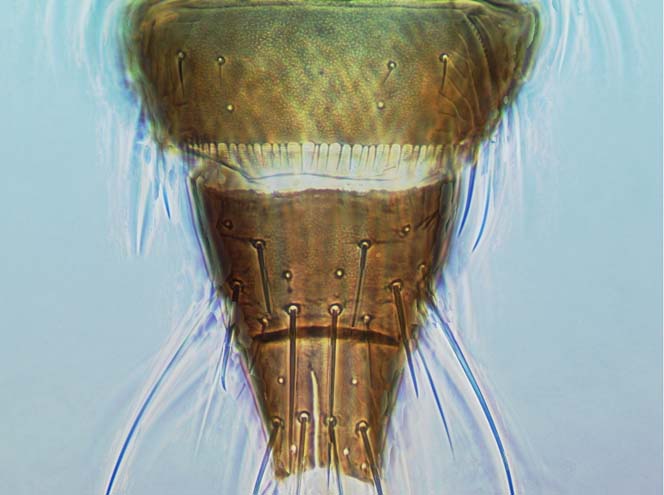
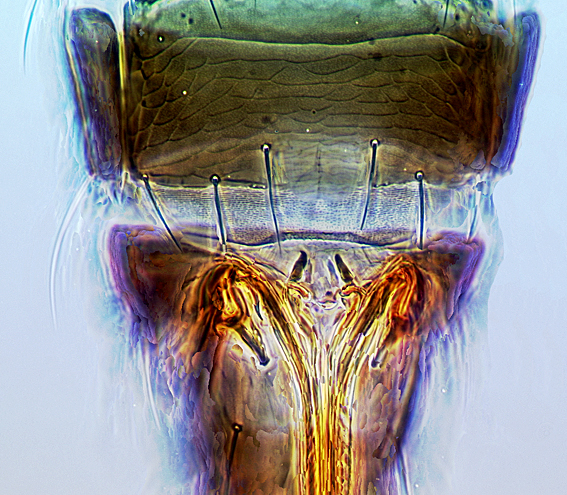
Most Thripidae have three pairs of setae associated with the ocelli, and the pronotum commonly bears two pairs of prominent posteroangular setae, but reduction in setal lengths on the pronotum and fore wings has occurred in several unrelated genera. The metanotal median pair of setae usually arise at the anterior margin, but are sometimes near the middle of this sclerite; they are close to the posterior margin only in the members of Synaptothrips from South Africa. The posteromarginal cilia on the fore wing are wavy (undulating) in most species of this family, and two longitudinal veins are usually visible.
The family Thripidae is currently interpreted as comprising rather more than 2000 described species, these being arranged into four subfamilies. The number of genera/species recognised in each of these subfamilies is as follows: Dendrothripinae 16/100; Panchaetothripinae 38/130; Sericothripinae 3/140; Thripinae 230/1700.
More than 40% of the genera of Thripidae are monobasic (ie. each one includes a single species), and scarcely 30 of the genera include more than 10 species. However, a few of these larger genera include more than 50 species, with genus Thrips including about 280, and Frankliniella about 220 species. Nomenclatural details are available on Mound’s Thysanoptera pages.
Thripidae species can usually be found wherever there is vegetation anywhere in the world. The majority of species occur in the tropics and warm temperate areas, but a few species are known from the subarctic (Greenland) and the subantarctic (Kerguelen and Macquarie Islands). In general, there is a considerable difference between both the size and composition of the thrips faunas of tropical and temperate areas, such as North/South across the land masses of both North America and Australia.
Some Thripidae species are particularly liable to be transported through human trading, such that some grass-living species (Aptinothrips rufus) are effectively worldwide (Mound, 1983). Similarly, Thripidae associated with particular crops are also widespread, such as the gladiolus thrips (Thrips simplex) and the onion thrips (Thrips tabaci). Some of these expanded distribution patterns probably occured more than 100 years ago, during the period of sailing ships, but the recent increase in air transport of horticultural produce is reflected in the increased distribution of other species, such as Scirtothrips dorsalis and Thrips palmi.
Europe is the only part of the world for which the Thripidae fauna can be considered reasonably well-known (zur Strassen, 2003), although even on that continent there is a lack of precise host-plant information for many common species. The Thripidae fauna of North America is probably less known than the available literature might imply (Stannard, 1968; Hoddle et al., 2008), and the fauna of Australia is certainly considerably larger than published figures indicate. Similarly, much of the Thripidae fauna of S.E. Asia remains undescribed, and the Thripidae fauna of tropical Africa is effectively unknown.
The Thripidae is one of eight families recognised in the Thysanoptera suborder Terebrantia.
The family Thripidae is currently interpreted as comprising rather more than 2000 species, these being arranged into four subfamilies. The Dendrothripinae is a group of small leaf-feeding thrips with an enlarged metathoracic furca that is associated with leg muscles involved in jumping. The Sericothripinae is a group of species found both in flowers and on leaves, and these (often bicoloured) thrips have the body surface bearing numerous rows of microtrichia, and the eighth antennal segment with a sensorium that has an exceptionally elongate, but slender, base. The Panchaetothripinae are largely tropical leaf-feeding thrips, in which the body and legs have strong reticulate sculpture. Thripinae is the largest group of species, and although most of them feed either on leaves or on flowers of higher plants, many being pests, a few species are predatory and a very few are associated with mosses or ferns.
Supra-generic groups within the Thripinae are difficult to define. Thrips genus-group and Frankliniella genus-group are both well-defined, and both comprise a large number of species in a series of genera (Mound, 2002). Similarly, genus-groups have been defined around Mycterothrips, Scirtothrips and Trichromothrips (Masumoto & Okajima, 2005, 2006, 2007). In contrast, some other groupings such as the “Aptinothripina” are based on character states such as “absence of long setae” that are evidently convergent, reduction in setal lengths having occured within genera in several unrelated lineages within the Thripidae.
Bhatti (2006) has presented a very different classification of these taxa in recognising three superfamilies, Dendrothripoidea (for Dendrothripinae plus Retithrips from the Panchaetothripinae); Rhipiphorothripoidea (for Rhipiphorothrips from the Panchaetothripinae), and Thripoidea (for the remaining taxa). This decision was based on character states of adults, but did not include larval characters. Larval Panchaetothripinae, including Retithrips and Rhipiphorothrips, have a characteritic body form with the tenth abdominal segment tubular, and their chaetotaxy is different from that of species in Dendrothripinae, Sericothripinae and Thripinae (Heming, 1991). These three superfamilies do not seem to represent evolutionary relationships.
Similarly, Bhatti (2006) recognised nine families within his “Thripoidea”, that is, eight families in addition to the Thripidae. Five of these eight families were further subdivisions of the Panchaetothripinae, one of them represents the Sericothripinae, one included a single genus specific to Pandanus in the Old World tropics, and one included a lineage of thrips that breed and pupate within individual grass flowers and thus have a distinctive body form. This classification is essentially phenetic, emphasising structural differences rather than relationships. It is thus of limited value to other biological disciplines, and therefore seems unlikely to be widely accepted (Mound & Morris 2007).